Mikrobielle Ökologie des Termitendarms
Forschungsbericht (importiert) 2003 - Max-Planck-Institut für terrestrische Mikrobiologie
Einleitung
Bei Insekten, die sich von verholztem Pflanzenmaterial ernähren, ist der Darm stark erweitert und beherbergt eine komplexe mikrobielle Lebensgemeinschaft. Vor allem die Termiten (Ordnung: Isoptera) haben als Paradebeispiel der symbiontischen Verdauung schon Generationen von Forschern fasziniert. Vieles deutet darauf hin, dass die Symbionten Stoffwechselleistungen bereitstellen, welche dem Wirt ansonsten nicht zur Verfügung stünden.
Meine Arbeitsgruppe beschäftigt sich schon seit einigen Jahren mit der Mikrobiologie und mikrobiellen Ökologie von Termitendärmen [1]. Unsere Untersuchungen zeigten, dass Insektendärme hervorragende Modellsysteme für die Bearbeitung grundsätzlicher Probleme der mikrobiellen Ökologie darstellen. Wir interessieren uns primär für die Grundlage der enormen Vielfalt mikrobieller Lebensgemeinschaften, den Zusammenhang zwischen Diversität und Stabilität, den Einfluss abiotischer Faktoren und biotischer Interaktionen auf die räumliche Organisation der Lebensgemeinschaft sowie die resultierenden Stoffflüsse im Ökosystem.
Termiten sind wegen ihrer weiten Verbreitung und enormen Biomasse, vor allem in tropischen Regenwäldern und Savannen, von globaler Bedeutung. Die Rolle von Termiten im Methanhaushalt der Atmosphäre wird schon seit Jahren diskutiert, jedoch ist der Zusammenhang zwischen Ernährungsweise und der Methanbildung durch die Darmsymbionten immer noch nicht verstanden. Auch die Rolle der Darmmikrobiota bodenfressender Termiten bei der Transformation und Mineralisierung der organischen Bodensubstanz ist von großem Interesse. Vor allem der von uns gefundene Umsatz stickstoffhaltiger Bodenbestandteile durch Termiten und andere humusfressende Insekten hat weitreichende Konsequenzen für die Stoffkreisläufe im Boden.
Der vorliegende Beitrag soll einen groben Überblick über die Thematik geben und auf die neueren Entwicklungen und offenen Fragen hinweisen. Für ausführlichere Informationen wird auf diverse Übersichtsartikel verwiesen [2-7].
Die symbiontische Verdauung
Die Hauptbestandteile pflanzlicher Zellwände sind Cellulose und Hemicellulosen. Die glykosidischen Bindungen zwischen den Untereinheiten dieser Polysaccharide sind im Prinzip leicht hydrolysierbar. Wegen der enormen Größe, den vielfältigen Primär- und Sekundärstrukturen sowie den physikalischen und chemischen Wechselwirkungen zwischen den einzelnen Komponenten erfordert aber bereits die Verdauung unverholzter Zellwände die synergistische Wirkung vieler verschiedener Enzyme. Da bei der Verholzung die Polysaccharide der Zellwände zudem in ein amorphes Netz phenolischer Polymere eingebettet werden, welches hauptsächlich nicht hydrolysierbare Bindungen aufweist und die Polysaccharide vor einem enzymatischen Angriff schützt, benötigt der Abbau verholzter Substrate außerdem noch einen Mechanismus zur Zerstörung des Lignocellulose-Komplexes.
, an die Darmwand angeheftet (b) oder Endobionten (c) und Epibionten (d) von Protozoen. Das Schema ist stark vereinfacht. (Verändert nach [4].)")

Cellulasen und andere Enzyme für den effizienten Abbau von Cellulose und Hemicellulosen findet man jedoch hauptsächlich bei Mikroorganismen. Pilze und bestimmte Bakterien (Actinomyceten) sind auch die einzigen Organismen, die eine Strategie entwickelt haben, das Lignin-Gerüst chemisch anzugreifen. Es ist darum nicht überraschend, dass sich Insekten und auch andere Tiere diese Kapazitäten zu Nutze machen und Mikroorganismen als Symbionten bei der Verdauung lignocellulosischer Nahrung einsetzen. Neben dem Pansen der Wiederkäuer ist der Enddarm von holzfressenden Termiten wohl das bekannteste Beispiel einer symbiontischen Verdauung.
Lignocellulose-Abbau durch Termiten
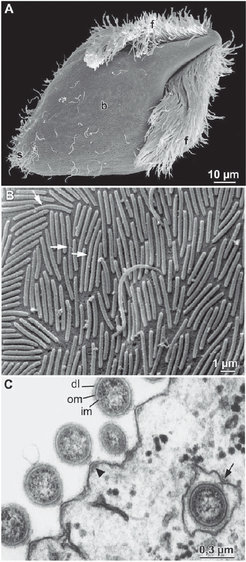
Am besten verstanden ist das Prinzip des Lignocellulose-Abbaus im Falle der holzfressenden niederen Termiten (Abb. 1). Neben einer großen Zahl von Prokaryonten beherbergt der erweiterte Enddarm dieser Tiere eine Vielfalt ungewöhnlicher Protozoen (Abb. 2). Diese sauerstoffempfindlichen Flagellaten sind essentiell für die Verdauung des Holzes. Sie phagozytieren die von der Termite zerkleinerten Holzpartikel und sind die Hauptquelle der cellulolytischen und hemicellulolytischen Enzymaktivitäten im Enddarm.
 Anaerobe Flagellaten in der Darmflüssigkeit (Phasenkontrastmikroskopie); (B) Prokaryonten in der Peripherie des Enddarms (Transmissionselektronenmikroskopie); und (C) die dichte mikrobielle Besiedelung der Darmwand (Rasterelektronenmikroskopie). (Mikrofotos aus [4].)")

Während der Endarm aller niederen Termiten mit Flagellaten vollgepackt ist, beherbergen die höheren Termiten (Familie: Termitidae) eine hauptsächlich prokaryontische Mikrobiota aus Bakterien und Archäen. Ursprünglich nahm man an, andere Mikroorganismen oder - im Falle der pilzkultivierenden Termiten - mit der Nahrung aufgenommene Cellulasen hätten die Funktion der cellulolytischen Protozoen übernommen. Neuere Untersuchungen zeigten jedoch, dass die cellulolytischen Aktivitäten in den vorderen Darmabschnitten aller Termiten von den Tieren selbst stammen. Das uralte Dogma, dass Tiere keine Cellulasen bilden können, hat sich spätestens mit dem Nachweis von Cellulase-Genen im Genom von Termiten und anderen Invertebraten und deren Expression in den Speicheldrüsen oder im Mitteldarmepithel endgültig überlebt [9].
Entgegen früherer Annahmen scheint der Ligninabbau im Termitendarm keine wichtige Rolle zu spielen. Vermutlich erzeugt die mechanische Zerstörung des Holzes durch Mandibeln und Kaumagen und die durch die Zerkleinerung bedingte Oberflächenvergrößerung ausreichend Angriffsfläche für Cellulasen und andere Enzyme. Lediglich in den Pilzgärten der Macrotermitinae verursacht der symbiontische Pilz einen weitgehend vollständigen Ligninabbau, und die höhere Effizienz des Lignocellulose-Abbaus durch pilzkultivierende Termiten deutet darauf hin, dass die kinetische Limitierung in Abwesenheit eines lignolytischen Systems nur teilweise aufgehoben wird.
Der Termitendarm als Bioreaktor
Im Gegensatz zu Insekten, die keine ausgeprägte Darmmikrobiota aufweisen, ist der Enddarm von Termiten deutlich verlängert und zu einer oder mehreren Kammern erweitert (Abb. 3). Obwohl man ursprünglich davon ausging, dass es sich dabei um reine Gärkammern handelt, erwies sich die Aufrechterhaltung anoxischer Verhältnisse in derart winzigen Systemen als keineswegs trivial. Unsere Untersuchungen mit Mikrosensoren zeigten, dass die steilen Konzentrationsgradienten zwischen oxischem Darmepithel und anoxischem Inhalt wegen des großen Oberflächen/Volumen-Verhältnisses einen enormen Sauerstoffeinstrom bedingen, dem nur der von den Gärungsprozessen getriebene Sauerstoffverbrauch durch die intestinale Mikrobiota entgegensteht.
")

Der vergrößerte Durchmesser des Enddarms dient folglich nicht nur der Verlängerung der Aufenthaltszeit der Nahrung im Intestinaltrakt, sondern verringert auch den Anteil des oxischen Darmvolumens. Erst dadurch wird Lebensraum für die sauerstoffempfindlichen Protozoen und Bakterien geschaffen und der unweigerliche Verlust von Gärprodukten durch oxidative Prozesse vermindert. Trotzdem hat der Einstrom von Sauerstoff in die Darmperipherie einen dramatischen Einfluss auf die Stoffwechselprozesse im intakten Darm. Viele der Bakterien und Protozoen im Termitendarm, vor allem diejenigen an der Darmwand, müssen besondere Anpassungen an die speziellen Sauerstoff-Verhältnisse ihres Lebensraums aufweisen. Isolate anaerober Bakterien aus Termitendärmen besitzen entsprechende Schutzenzyme und können selber mit hoher Rate Sauerstoff reduzieren.
Die Akkumulation von Gärprodukten, die unerwartet steilen Wasserstoffgradienten, und die Bildung von Methan belegen jedoch, dass der Enddarm trotz allem eine anaerobe Mikrobiota beherbergt. Das ursprüngliche Konzept des Stoffwechsels im Darm niederer Termiten ging davon aus, dass die anaeroben Flagellaten die Polysaccharide hydrolysieren und dann zu Acetat, Wasserstoff und CO2 vergären. Durch Mikroinjektion von Radiotracern in intakte Därme konnte jedoch gezeigt werden, dass das Spektrum der von den Protozoen gebildeten Gärprodukte weitaus größer sein muss als angenommen, und das vor allem Laktat ein Schlüsselintermediat in der mikrobiellen Nahrungskette des Enddarms darstellt.
Die Gärprodukte der Protozoen bilden neben den direkt aus dem Mitteldarm stammenden Metaboliten die Grundlage für die Existenz unzähliger Bakterien, welche in der Darmflüssigkeit leben, an der Darmwand angeheftet sind oder Cytoplasma und Zelloberfläche der Protozoen besiedeln (Abb. 4 und 5). Die von den mikrobiellen Gärungen gebildeten kurzkettigen Fettsäuren (vor allem Acetat, Propionat, und Butyrat), welche letztendlich in der Enddarmflüssigkeit akkumulieren, werden vom Darmepithel resorbiert und dienen der Termite als Substrat.
 Gefrierschnitt des Enddarms einer holzfressenden Termite (unten rechts), dessen Wand mit einem Biofilm von Bakterien (rote Fluoreszenz) belegt ist. Die Kerne der Epithelzellen und der Protozoen im Enddarm erscheinen blau (Mikrofoto: D. Schmitt-Wagner). Viele der Wasserstoff produzierenden Protozoen (B) sind eng mit Wasserstoff verbrauchenden methanogenen Archäen vergesellschaftet, die an ihrer grünen Eigenfluoreszenz zu erkennen sind (C) (Mikrofotos: M. Pester).")

Struktur-Funktionsbeziehungen in der Darmmikrobiota
Die phylogenetische Charakterisierung der prokaryontischen Mikrobiota von Termitendärmen, welche erst durch die molekularen, kultivierungsunabhängigen Ansätze der letzten Jahre ermöglicht wurden, dokumentiert eine enorme phylogenetische Diversität. Es ist auffällig, dass viele der Symbionten Entwicklungslinien vertreten, die ausschließlich im Darm von Termiten und anderen Insekten vorkommen. Vor allem die Epibionten und Endobionten der Protozoen sind oft nur entfernt mit bekannten Arten verwandt, und die Aufklärung ihres Stoffwechsels dürfte den Schlüssel zum Verständnis dieser Symbiosen darstellen. Die Untersuchungen stehen jedoch noch am Anfang, und wie in anderen Bereichen der mikrobiellen Ökologie ist es eine der vordringlichsten Aufgaben, den einzelnen Vertretern der mikrobiellen Lebensgemeinschaft bestimmte Funktionen zuzuweisen und die an den wesentlichen Stoffwechselprozessen beteiligten Populationen zu identifizieren und zu lokalisieren.
Besonders interessant ist diese Frage im Falle des intermediär gebildeten Wasserstoffs, an dessen Oxidation wenigsten zwei metabolisch grundverschiedene Gruppen beteiligt sind: Methanogene Archäen, welche CO2 zu Methan reduzieren, und homoacetogene Bakterien, welche CO2 zu Acetat reduzieren. Methanogenese und reduktive Acetogenese kommen prinzipiell in den Enddärmen aller Termiten vor. In den meisten holzfressenden Arten übertrifft jedoch reduktive Acetogenese die Methanogenese als Wasserstoffsenke, wodurch der für den Wirt verfügbare Pool von kurzkettigen Fettsäuren substantiell erhöht wird. Obwohl eine alternative Produktion von Methan für den Wirt kaum von Vorteil sein kann, ist die Methanogenese bei pilzkultivierenden und bodenfressenden Termiten erheblich stärker. Ein Verständnis der grundsätzlichen Zusammenhänge wäre wünschenswert, hat doch die Methanemission dieser Termitengruppen einen signifikanten Anteil am globalen Budget dieses wichtigen Treibhausgases.
 zahlreiche stäbchenförmige Bakterien (b) und einige Spirochäten (s) (A, B). Dünnschnitt der epibiontischen Stäbchen (C). (Elektronenmikroskopische Aufnahmen aus [8].)")
Problem Stickstoffhaushalt
Lignocellulose ist nicht nur ein schwer abbaubares, sondern auch ein extrem nährstoffarmes Substrat. Das C/N-Verhältnis im Holz ist mehr als hundertfach höher als das des Insektenkörpers. Außerdem fehlen einer lignocellulosischen Diät die meisten der von der Termite benötigten essentiellen Nährstoffe, wie Aminosäuren, Vitamine und Sterole. Viele Mikroorganismen sind jedoch in der Lage, Stickstoff zu fixieren, Nitrat und Ammonium zu assimilieren und die von den Tieren benötigten Aminosäuren und Vitamine zu synthetisieren.
Im Fall der pilzkultivierenden höheren Termiten (Unterfamilie: Macrotermitinae), welche Pilze der Gattung Termitomyces (Basidiomyceten) innerhalb des Nestes in so genannten Pilzgärten auf vorverdauter Nahrung züchten, wandelt der symbiontische Pilz einen Teil der Lignocellulose in ernährungsphysiologisch wertvollere pilzliche Biomasse um, die anschließend verdaut wird. Da der Inhalt des Enddarms nicht direkt verdaut werden kann, müssen alle anderen Termiten flüssigen Darminhalt von den Nestgenossen erbetteln. Diese proctodeale Trophallaxis (Abb. 1) ist ein Sozialverhalten, welches auch der Weitergabe der Symbionten dient und nur bei Termiten vorkommt. In den letzten Jahren häufen sich die Hinweise, dass im Speichel und im Mitteldarm von Termiten alle wesentlichen Enzyme zur Verdauung von pilzlicher und mikrobieller Biomasse bereitgestellt werden.
Wie bei allen Insekten werden Harnsäure und Harnstoff als Abfallprodukte des tierischen Nukleinsäure- und Proteinabbaus über die Malpighischen Gefäße in den Enddarm abgegeben. Die Mineralisierung dieser Substanzen durch die Mikrobiota des Enddarms und die anschließende Assimilation des Ammoniums in mikrobielle Biomasse sorgt dafür, dass Termiten keinen Stickstoff mit dem Kot verlieren. Die hohen Ammoniumkonzentrationen im Darm erlauben ein gutes Wachstum der Mikroorganismen - quasi eine Stickstoffdüngung der intestinalen Mikrobiota. Der Pool an verfügbarem Stickstoff wird außerdem durch die Aktivität Stickstoff-fixierender Bakterien ständig vergrößert. Man schätzt, dass etwa die Hälfte des Stickstoffs im Gewebe von Trockenholz-Termiten der Atmosphäre entstammt.
Bodenfressende Termiten
Während die niederen Termiten noch fast ausnahmslos Holz fressen, zeigen die evolutionär weiter entwickelten höheren Termiten (Familie: Termitidae) eine Diversifizierung in der Ernährung. Die bereits erwähnten pilzkultivierenden Macrotermitinae, welche auf den Abbau von stickstoffarmer Pflanzenstreu spezialisiert sind, profitieren neben den Cellulose- und Lignin-abbauenden Aktivitäten vermutlich auch von der Fähigkeit des Pilzes zur Ammonium-Assimilation. Das Nahrungsspektrum der anderen Unterfamilien (Nasutitermitinae, Termitinae und Apicotermitinae) reicht von totem Holz über verholztes Pflanzenmaterial in verschiedenen Stadien der Humifizierung bis hin zu Humus und Tierdung. Mit zunehmender Humifizierung steigt das C/N-Verhältnis in der Nahrung, und die im Substrat enthaltene mikrobielle Biomasse gewinnt an Bedeutung.
Über die Ernährungsgrundlage der bodenfressenden Termiten, welche in tropischen Ökosystemen eine wichtige Rolle spielen, war bis vor kurzem so gut wie nichts bekannt. Die Därme dieser Tiere sind besonders stark verlängert und in mehrere aufeinanderfolgende Kammern gegliedert (Abb. 6), von denen jede andere physikochemische Verhältnisse aufweist und eine spezielle Mikrobiota beherbergt. Unsere Untersuchungen belegten einen eindeutigen Zusammenhang zwischen dem extrem alkalischen pH im vorderen Enddarm dieser Tiere, welcher sogar pH 12 übersteigen kann, und der Spezialisierung auf die prinzipiell nährstoffreiche, aber schwer zu erschließende Diät. Erst die alkalische Extraktion aus der Bodenmatrix ermöglicht einen enzymatischen Angriff auf die organische Substanz, welche ansonsten vor dem mikrobiellen Abbau im Boden geschützt sind.
 spielen eine wichtige Rolle im Stoffkreislauf tropischer Ökosysteme. Der Darm der Arbeiter ist mit Boden gefüllt und gegenüber den Därmen holzfressender Termiten verlängert und stärker kompartimentiert (B). Das erste Enddarmsegment (P1) ist extrem alkalisch (pH > 12). (Verändert nach [7].)")

Partner oder nur Mitesser?
Der Tatbestand der mutualistischen Symbiose erfordert einen gegenseitigen Vorteil der beteiligten Partner. Der Darm bietet der Darmmikrobiota hervorragende Lebensbedingungen; der Wirt sorgt für eine kontinuierliche Versorgung mit Substraten und sichert über die Weitergabe der Mikrobiota an Nestgenossen auch deren Verbreitung. Im Gegenzug ist auch der Nutzen der Cellulose-abbauenden Flagellaten im Darm niederer Termiten nachvollziehbar und experimentell belegt. Bei vielen prokaryontischen Symbionten hingegen ist der Nutzen nicht offensichtlich, und in manchen Fällen scheint ihre Aktivität sogar von Nachteil. Damit wären sie lediglich Kommensalen und der Wirt könnte von ihrer Entfernung eigentlich nur profitieren.
Allerdings ist der Energieverlust an die Darmmikrobiota unter sauerstofflimitierten Bedingungen nur gering. So setzt etwa ein homoacetogener Abbau nur etwas mehr als 10% der in der Cellulose enthaltenen biologisch nutzbaren Energie um - der Rest steht dem Wirt in Form der Gärprodukte weiterhin zur Verfügung. Mögliche Vorteile wie Stickstoff-Fixierung, Ammonium-Assimilation, Versorgung mit Vitaminen oder auch die Verhinderung einer Besiedelung des Darms durch potenziell pathogene Mikroorganismen in Anwesenheit einer stabilen Mikrobiota könnten diesen Einsatz durchaus rechtfertigen.